|
||||
|
|
Глава шестая РЕАКЦИИ НЕРВНОЙ СИСТЕМЫ СОБАК В УСЛОВИЯХ ЭКСТРЕМАЛЬНЫХ ФАКТОРОВ Известно, что центральная нервная система играет ведущую роль как высший интегрирующий орган и ее функциональное состояние имеет решающее значение для общего состояния живых организмов. Поэтому тщательное описание и анализ функций нервной системы в условиях воздействия экстремальных факторов являются актуальной задачей современного этапа развития биологии. Расширение задач, стоящих перед космической физиологией и медициной, существенно повышает значение этой проблемы. Их разработке также способствует необходимость перехода к длительным космическим полетам, к организации жизни орбитальных станций и т. д. Научные исследования советских ученых в течение ряда лет были направлены на изучение физиологических функций организма собак и, в частности, функций центральной нервной системы при действии на нее различных экстремальных факторов, возникающих при космических полетах. Условно-рефлекторная деятельность при экстремальных воздействиях изучалась, начиная с первых шагов развития космической биологии и медицины, под руководством профессоров О.Г. Газенко, Н.Н. Лившиц, Н.А. Рокотовой (Лукьянова, 1962; Рокотова и др., 1962; Васильев, Герд, 1964; и др.). Начиная с 1961 г., внимание исследователей стали привлекать биоэлектрические реакции структур, связанных с мозгом. Методом электроэнцефалографии была зарегистрирована высокая биоэлектрическая активность в гипокампе и миндалевидном ядре мозга кошек в период воздействия ускорений. Активность при увеличении перегрузок до +5–6g на 10–15 сек. превращалась в судорожные разряды, распространяющиеся на подкорку (Adey et al., 1961). Изучалась (Изосимов, Разумеев, 1962 a, b; Газенко и др., 1964) биоэлектрическая активность ретикулярной формации и коры, а также их взаимосвязь. В отдельных нейронах гигантоклеточного ядра ретикулярной формации кошек при действии ускорений в первом периоде регистрировалось учащение импульсации, во втором — залповидная импульсация и потом (в третьем периоде) импульсация отсутствовала. Ретикулярная формация и гипоталамус, по мнению авторов, быстро реагировали на действие ускорений и влияли на кору мозга. Торможение, развивающееся в ретикулярной формации, предохраняло кору от повреждающегося действия ускорений (Газенко и др. 1963). Вначале это были работы, направленные на изучение влияния на условно-рефлекторную деятельность отдельно взятых перегрузок, вибраций, шумов, облучений (Михеева, 1955; Кисляков, 1956, 1958; Скачедуб, 1957; Павлова, 1958; Гуровский, 1959; Росин и др., 1959; Барер, 1962 а, б; Борщевский и др., 1963; Кузнецова, 1964; Лившиц, Мейзеров, 1967; Климовицкий, 1967; и др.). Однако в ходе космического полета на животных и человека действуют не отдельные факторы его, а целый ряд одновременных или последовательных воздействий. Реакции на такое комбинированное воздействие особенно сложны и многообразны: можно ожидать, что они не будут соответствовать таковым при изолированном действии отдельных факторов. В связи с этим изучение интегрального ответа на комплексы раздражения должно было представлять большой теоретический и практический интерес. Возник вопрос о возможности эффектов частичной суммации, взаимного ослабления или нейтрализации реакции при различных формах комплексного воздействия. Практический план исследований означал подготовку к выработке ряда рекомендаций, направленных на уменьшение силы отрицательно воздействующих условий при будущих космических полетах. С переходом к изучению таких интегрально-действующих факторов космического полета и связан новый, второй этап исследования. Различные формы комплексного воздействия на нервную систему успешно изучались целым рядом исследователей. Особенно интересны и многочисленны работы, выполненные по этой тематике под руководством профессора Н.Н. Лившиц. Большинство подобных исследований посвящено влиянию на реакции нервной системы сразу двух факторов (Зуев, 1960; Кузнецова, 1964; Лившиц, Мейзеров, 1966; Кузнецова, 1966; Апанасенко, 1966; Кузнецова, 1967; Лившиц, Мейзеров, 1967; и др.). Полученные данные убедительно показали, что воздействие на нервную систему двухчленного комплекса качественно отличалось от воздействия каждого отдельно взятого фактора. Очень важно, что в ряде случаев возникал эффект ослабления отрицательно действующих раздражений, например, вибрации повышали устойчивость организма к облучению. Несмотря на то что в настоящее время наметился переход к изучению комплексного влияния факторов космических полетов, науку также продолжают интересовать исследования, направленные на углубленное изучение воздействий отдельно взятых факторов. Более того, проведенные исследования (Лившиц, 1967) показали, что интегральный ответ нервной системы на комплекс раздражений может быть изучен и правильно осмыслен только в том случае, если ему предшествовали исследования влияния на живые организмы отдельно взятых факторов. Описанные эксперименты имели цель исследовать изменение реакций центральной нервной системы животных на отдельно взятые вибрации, перегрузки, а также в период космического полета. Опыты проводились на собаках, так как эти животные использовались при запусках ракет и спутников. Работа с собаками имела свои преимущества: они легко привыкали к экспериментальной обстановке, спокойно вели себя в опытах, нормальная физиология и условнорефлекторная деятельность этих животных хорошо изучены. Эксперименты проводились в 1958–1962 гг. Исследование влияния на реакции центральной нервной системы собак ускорения, вибрации и космических полетов осуществлялось с помощью четырех серий опытов; влияние на условнорефлекторную деятельность собак перегрузок и вибраций изучалось на 16 собаках, космических полетов — на трех собаках. Кроме того, проводились эксперименты по изучению изменений после полета поведенческих реакций и психического состояния трех собак. Определение особенностей условнорефлекторной деятельности собак Исследование высшей нервной деятельности собак во всех сериях опытов проводилось с помощью методики и аппарата, описанных в начале книги. Создавая методику, мы исходили из того, что она должна была выявлять наиболее тонкие изменения в высшей нервной деятельности. Поэтому в качестве индикатора использовались сенсомоторные реакции собак, близкие по своему выражению к двигательным пищедобывательным (Воронин, 1957). Вырабатываемые рефлексы имели сложную эффекторную часть, требующую тонкой координации. Поэтому они были более подвержены изменениям, чем слюнные, электрокожные и кислотные. По своему выражению и процессу выработки получаемые рефлексы были ближе всего к так называемым произвольным движениям. По нашему мнению, это должно было способствовать выявлению у животных в период воздействия на них экстремальных факторов скрытых тонких изменений в реакциях центральной нервной системы. Изучению рефлекторной деятельности предшествовало исследование у подопытных собак особенностей реакций их центральной нервной системы. Известно, что определение типа собак даже по малому стандарту испытаний в школе И.П. Павлова проходило на протяжении многих месяцев и включало в себя изучение процесса выработки условных рефлексов, исследование угашения с подкреплением и без подкрепления, переделку сигнальных раздражителей, пробы с применением фармакологических средств и т. д.  Рис. 22. Обследование высшей нервной деятельности путем трехкратной подачи выработанного стереотипа раздражителей Собака Стрелка (2. VI 1960 г.); а, б — рефлексы на положительные сигналы Рис. 22. Обследование высшей нервной деятельности путем трехкратной подачи выработанного стереотипа раздражителей Собака Стрелка (2. VI 1960 г.); а, б — рефлексы на положительные сигналы В наших условиях использование существующих лабораторных стандартов для определения типов было не применимо по ряду причин. Использование дрессировки лишало возможности охарактеризовать процесс выработки условных рефлексов. Угашение с подкреплением и без подкрепления и т. д. мы не могли применить, так как в каждый момент необходимо было иметь хорошо закрепленные реакции на стереотип раздражителей и т. д. В литературе (Паршутин и др., 1954; Флес, 1958) описывались приемы более простой и доступной оценки типологических особенностей животных. Но эти методы также не могли быть применены в наших условиях. В связи со сказанным нам пришлось на основании принципов павловских функциональных проб продумать свою систему проверки особенностей высшей нервной деятельности собак. Оценочная роль такой системы не могла быть детально изучена. Поэтому целью исследований было охарактеризовать, хотя бы частично, процессы возбуждения, торможения и их баланса. Это позволяло сделать обоснованное заключение об отсутствии у подопытных животных патологических черт реакций центральной нервной системы. Обследование нервной деятельности животных включало трехкратную подачу выработанного стереотипа, пробу для определения силы возбудительного процесса, две пробы для определения тормозного процесса и две — для испытания баланса возбудительных и тормозных процессов.  Рис. 23. Обследование процесса возбуждения путем подачи положительных раздражителей через укороченный в два раза интервал, т. е. через 15 сек. А — Белка (10. V 1960 г.); Б — Звездочка (16. I 1960 г.); в — межсигнальные реакции Рис. 23. Обследование процесса возбуждения путем подачи положительных раздражителей через укороченный в два раза интервал, т. е. через 15 сек. А — Белка (10. V 1960 г.); Б — Звездочка (16. I 1960 г.); в — межсигнальные реакции Трехкратная с интервалами в 10–17 мин. подача выработанного стереотипа раздражителей позволяла точнее оценить условнорефлекторный фон и провести статистическую обработку некоторых показателей реакции центральной нервной системы. У собаки Стрелки, отобранной для космического полета, такая подача раздражителей не вызывала нарушений условнорефлекторной деятельности, как это видно из рис. 22. На рис. 22–26 показана условнорефлекторная деятельность подопытных собак при перечисленных выше пробах. Действие положительных сигналов обозначалось буквой «К», тормозных — «Ж» и «3» (свет большой и средней интенсивности), рефлексы на положительный раздражитель — точкой, на тормозной и срывы дифференцировок — кружочком, обведенным двумя линиями. Межсигнальные реакции обозначались кружочком, обведенным одной линией. Расположение этих обозначений по оси ординат примерно соответствует той секунде, на которой появлялись данные реакции. Для определения силы возбудительного процесса — способности испытывать сильное возбуждение — использовалось многократное (20-разовое) повторение положительных сигналов при уменьшении интервалов между раздражителями (интервал в 30 сек. заменялся интервалом в 15 сек.). Проведение таких проб показано на рис. 23. Считалось, что животные обладают сильным возбудительным процессом, если уменьшение интервалов не сопровождалось запредельным торможением, существенным изменением латентного периода, появлением большого числа межсигнальных реакций.  Рис. 24. Испытание тормозного процесса с помощью удлинения (до 15–25 сек.) дифференцированного тормозного раздражителя А — Белка (7. V 1660 г.); Б — Жулька (18. Х 1960 г.). Остальные обозначения те же, что на рис. 22, 23 Рис. 24. Испытание тормозного процесса с помощью удлинения (до 15–25 сек.) дифференцированного тормозного раздражителя А — Белка (7. V 1660 г.); Б — Жулька (18. Х 1960 г.). Остальные обозначения те же, что на рис. 22, 23 Сила тормозного процесса — способность выдерживать длительное концентрированное торможение — испытывалась с помощью традиционной для павловской школы пробы с удлинением до 15 и 25 сек. (вместо 10 сек.) действия тормозного раздражителя (рис. 24). В этом варианте раздражители подавались в том же порядке, что и обычно. Первые две дифференцировки — свет большой и средней интенсивности — удлинялись до 15 сек., а вторые две — до 20 сек. С целью охарактеризовать процесс торможения также применялась показанная на рис. 25 проба с удлинением до 60 сек. (вместо 30 сек.) в обычном стереотипе интервала перед тормозным раздражителем.  Рис. 25. Испытание тормозного процесса с помощью удлинения (до 60 сек.) интервала перед тормозным раздражителем А — Жулька (18. Х 1960 г.); В — Звездочка (14. I 1961 г.). Остальные обозначения те же, что на рис. 22, 23 Рис. 25. Испытание тормозного процесса с помощью удлинения (до 60 сек.) интервала перед тормозным раздражителем А — Жулька (18. Х 1960 г.); В — Звездочка (14. I 1961 г.). Остальные обозначения те же, что на рис. 22, 23 Для испытания баланса возбудительного и тормозного процессов использовалась проба с перемежающейся подачей положительных и тормозных раздражителей (рис. 26). В качестве дифференцировок, чтобы исключить сильно действующий свет, применялся только свет средней интенсивности. 25 раздражителей подавалось в следующем порядке: три положительных и дифференцировка, затем один положительный и дифференцировка, потом два положительных и дифференцировка, вслед за этим два раза подряд один положительный и дифференцировка, затем после положительного две дифференцировки, потом опять положительный и дифференцировка и 2 раза подряд положительный и две дифференцировки. Заканчивалась система подаваемых сигналов положительным раздражителем. Результаты обследования с помощью этой пробы показаны на рис. 26.  Рис. 26. Испытание баланса возбудительного и тормозного процессов путем перемежающейся подачи дифференцировочных и положительных раздражителей А — Звездочка (13. II 1961 г.); Б — Жулька (15. VIII 1966 г.); г — срывы дифференцировки. Остальные обозначения те же, что на рис. 22, 23 Рис. 26. Испытание баланса возбудительного и тормозного процессов путем перемежающейся подачи дифференцировочных и положительных раздражителей А — Звездочка (13. II 1961 г.); Б — Жулька (15. VIII 1966 г.); г — срывы дифференцировки. Остальные обозначения те же, что на рис. 22, 23 Кроме приема с перемеживающейся подачей положительных и тормозных раздражителей, для испытания баланса нервных процессов применялась также проба с увеличенным интервалом (45 сек. вместо 30 сек.) Раздражители следовали в том же порядке, как и в обычном стереотипе. Увеличенные интервалы предваряли 3, 10 и 13–15-й раздражители. Перед 11-м раздражителем (тормозным большей силы) интервал был нормальным. Опыты проводились на собаках, отобранных по характеру их поведения и высшей нервной деятельности. Брали животных, которые в клетках вивария и на прогулках в двигательном и голосовом отношениях вели себя спокойно. Исследование влияния на реакции центральной нервной системы перегрузок, вибраций, шума, космического полета проводилось на 12 собаках основной группы, выбранных из 37 животных; 25 животных было выбраковано в связи с тем, что у них при испытании особенностей условнорефлекторной деятельности были обнаружены случаи выпадений реакций в ответ на положительные сигналы, срывов дифференцировок, большие вариации в латентном периоде и т. д. В группу собак, отобранных для запуска в космос, вошли животные, условнорефлекторная деятельность которых заслуживала наиболее высоких оценок не только во время проб на возбудительный и тормозной процесс, но также и при испытании баланса возбудительного и тормозного процессов. На основании полученных фактов можно было высказать предположение, что, например, собака Белка относилась к числу животных с сильными и уравновешенными нервными процессами. Данные, полученные при троекратном обследовании выработанного стереотипа у всех 12 собак, показали общую устойчивость условнорефлекторного фона. Собаки реагировали на 96% подаваемых раздражителей. Латентный период в среднем был равен 1,4 при сигме 0,5. Сила нажима на педаль и направленность движений отвечали предъявляемым к ним требованиям в 99,6%. Неправильные нажимы (у Малька и Быстрого) были единичными случаями. У большинства собак дифференцировки были абсолютными. Испытание возбудительного процесса показало, что собаки реагировали на 87% положительных сигналов. У семи из них было 100% правильных ответов, у пяти отмечалось выпадение небольшого числа реакций. Латентный период не изменялся или незначительно колебался, уменьшаясь и увеличиваясь (в среднем на 0,571 сек.). Укорочение интервала приводило к тому, что у половины собак межсигнальных реакций не было, у остальных они возникали в относительно небольшом числе. При испытании тормозного процесса было сохранено 68,7% тормозных реакций. У трех собак они были абсолютными, остальные правильно реагировали на три, два и один тормозной раздражитель. У всех собак наблюдалось большое количество (в среднем 12) межсигнальных реакций. Такие реакции чаще появлялись после дифференцировок и особенно в том случае, когда два тормозных раздражителя следовали друг за другом. Удлинение дифференцировок незначительно влияло на положительные реакции: отмечалось выпадение 7,5% ответов на сигналы. Скрытый период реакций несущественно колебался. Примерно такие же результаты были получены при испытании тормозного процесса с помощью другой пробы. Было сохранено 66,6% дифференцировок, т. е. всего на 2,1% меньше, чем при предыдущей. Регистрировалось выпадение 6% реакций на положительные и 27% на тормозные сигналы. Большое количество межсигнальных реакций наблюдалось в периоды удлиненных интервалов между отрицательными раздражителями и особенно, когда друг за другом следовало два тормозных сигнала. Абсолютные дифференцировки и реакции на все положительные раздражители имели место у трех животных, у других трех они также были весьма благоприятными: собаки реагировали на 10 из 11 положительных сигналов и на 3–4 отрицательных из четырех. У следующих трех положительные реакции нарушались относительно мало, а дифференцировки — существенно. У Звездочки, наоборот, при полной сохранности реакций на дифференцировочные раздражители отмечалось выпадение реакций на положительный сигнал. Все это позволяло сделать не только обоснованный вывод об отсутствии у подопытных животных патологических изменений в реакциях центральной нервной системы, но также и отнести Лисичку, Быстрого, Белку, Каму к собакам с сильными и уравновешенными процессами возбуждения и торможения. В нервной системе Невы, Малька и Пушинки, очевидно, при сильном раздражительном процессе обнаруживалась некоторая слабость тормозного. Она могла быть как результатом большей силы возбудительного процесса, так и слабости тормозного. Изменения латентного периода и число межсигнальных реакций в различных функциональных пробах давали возможность предположить у Невы первое, а у двух других животных — второе. Влияние ускорений на высшую нервную деятельность собак Влияние ускорений на реакции центральной нервной системы изучались целым рядом отечественных и зарубежных исследователей. Опубликованные данные были обобщены в нескольких обзорах (Chambers, 1963; Климовицкий, Лившиц, Родионов, 1967; и др.). Полученный материал говорил о существенных изменениях под влиянием ускорений психических реакций животных. Ускорения +0,5g в течение 3 мин. снижали процент правильных реакций крыс (Herrick et al., 1958). Процент снижения таких реакций был существеннее при ускорении +2g, действующем от 1 до 5 мин. (De Marko, Geller, 1964). При поперечных ускорениях вида +2g в течение 30 мин. условный оборонительный рефлекс шимпанзе, когда нажим на рычаг прекращал болевое раздражение, увеличивался и, наоборот, уменьшался, когда ускорение становилось большим (+4g). У Барера (1962а, б) ускорения вида +4g в течение 15 сек. вызывали нарушения условных рефлексов, а когда величина ускорений достигала +7g и +10g в течение 1 мин., наблюдался полный срыв условнорефлекторной деятельности. Для ответа на вопрос о глубине изменений реакций центральной нервной системы Барер (1962а, б) воспользовался методом исследования процесса восстановления. Восстановление условнорефлекторной деятельности крыс при действии на них ускорением вида +7g началось через 10–15 мин., а при +10g через 22–30 мин. Полностью животные отреагировали после ускорений вида +10g на подаваемый стереотип раздражителей только через 60 мин. Все это позволяло связать степень разрушения условнорефлекторной деятельности с силой воздействия ускорений. Такой вывод делал необходимым изучение реакций центральной нервной системы животных при величинах ускорений, ожидаемых в космическом полете. Н.Н. Лившиц и Е.С. Мейзеров (1967) подробно исследовали характер нарушений реакций центральной нервной системы у крыс. Они установили, что нарушения заключаются в выпадении некоторых рефлекторных ответов на положительные раздражители, в удлинении латентного периода условных рефлексов, в снижении силы реакций на положительные раздражители. Некоторые из этих нарушений также были характерны для высшей нервной деятельности собак: снижались положительные условные рефлексы, нарушались индукционные отношения и т. д. (Кисляков, 1956). В некоторых исследованиях были показаны резкие изменения рефлекторной деятельности в зависимости от характера подкрепляющих воздействий: болевых, кислотных, пищевых, питьевых (Herrick, 1961; и др.). Очевидно, разрушаемость и сохранность рефлексы, нарушались индукционные отношения и т. д. О механизмах, лежащих в основе нарушений реакций центральной нервной системы под действием ускорений, можно думать, что они создавали предпосылки для повышения возбудимости нервной системы. Она проявлялась в таких неспецифических реакциях, как учащение дыхания, появление тахикардии. В электроэнцефалограмме регистрировалась десинхронизация, активация ?-ритма. Кроме того, усиливалась реакция экзальтации ?-ритма на закрывание глаз, увеличивалась амплитуда вызванных потенциалов в зрительной коре при стимуляции глаза мелькающим светом (Барер, 1962а, б; Изосимов, Разумеев, 1962а, б; Росин и др. 1959; и др.). Усиление возбудимости также доказывалось увеличением лабильности и проводимости рефлекторных дуг, что выражалось в значительных изменениях латентного периода условных рефлексов при относительно постоянной или несущественно изменяемой величине рефлексов. После периода повышения и ослабления возбудимости понижается не только возбудительный, но и тормозной процесс. Об этом свидетельствовало нарушение нормальных индукционных отношений (Кисляков, 1956), а также случаи ультрапарадоксальных реакций (Лившиц, Мейзеров, 1967). Нарушения возбудительного и активного тормозного процесса приводили к преобладанию запредельного тормозного. Развивалось состояние разлитого торможения, вызывающее иногда сон (Васильев, Герд, 1964). В некоторых опытах наблюдался другой порядок развития всех этих явлений: условные рефлексы на первые раздражители отсутствовали, потом оказывались нормальными, в конце опыта опять регистрировалось их выпадение — циркуляторные нарушения нервной деятельности. Можно говорить о наличии определенного параллелизма между изменениями гемодинамических показателей и некоторыми функциональными показателями реакций нервной системы. Значительные нарушения возникали в случаях, если направление ускорения совпадало с направлением основных кровеносных сосудов. Характеризуя общее действие ускорений на нервную систему, профессор Н.Н. Лившиц указывала, что ускорения вызывали резкие, но кратковременные сдвиги. Дальнейшие исследования были проведены в плане отбора и подготовки собак к запускам в космос и входили как составная часть в более широкую группу изучения собак при действии на них ускорений (Котовская, Юганов, 1962). Центростремительные ускорения переменной величины до +10g создавались на центрифуге радиусом 3,7 м и действовали в поперечном направлении к оси тела грудь — спина. Величины ускорений, скорость их нарастания и время действия воспроизводились соответственно графикам выведения космических кораблей на орбиту. Опыты с воздействием перегрузок чередовались с контрольными, в которых сохранялись все условия эксперимента, за исключением вращения животных. Изучению реакций центральной нервной системы в связи с воздействиями ускорений предшествовало приучение животных находиться в фиксированном состоянии в кабине на центрифуге. Систему выработанных условных рефлексов проверяли непосредственно за 10–15 мин. до воздействия ускорений, через 5–7 мин. и через 25 мин. после центрифугирования. На рис. 27 показана собака Кама в кабине центрифуги. Полученные данные показали, что при обследовании высшей нервной деятельности через 5–7 мин. после воздействия глубокие нарушения были обнаружены у трех собак: резкое удлинение скрытого периода рефлексов, выпадение большого числа ответных реакций. Собаки Малек и Звездочка отреагировали более чем на половину подаваемых сигналов. При этом латентный период у них был замедлен относительно несущественно. Одно животное вначале подачи стереотипа раздражителей реагировало гораздо быстрее, чем обычно, а с пятого положительного сигнала, наоборот, его реакции удлинялись. Менее всего центрифугирование повлияло на количество ответных реакций и величину латентного периода у Невы: собака не отреагировала только на два сигнала, следующих за дифференцировкой. Однако у нее оказались значительно нарушенными дифференцировки. То же самое было характерно и еще для четырех собак. Таким образом, полученный материал показал, что не только у разных собак, но и у одного и того же животного по одним показателям реакций центральной нервной системы регистрировалось явное и резкое ухудшение условнорефлекторной деятельности, а по другим, наоборот, реакции были нормальными (табл. 15). Следовательно, у собак под влиянием ускорения в одних случаях нарушался процесс возбуждения (выпадение рефлексов в ответ на положительный раздражитель, замедление латентного периода), в других — процесс торможения (срывы дифференцировок, появление большого числа межсигнальных реакций и т. д.).  Рис. 27. Собака Кама в кабине центрифуги Собака одета в одежду, предохраняющую электроды для записи физиологических функций, и привязана для создания особым образом направленных перегрузок Рис. 27. Собака Кама в кабине центрифуги Собака одета в одежду, предохраняющую электроды для записи физиологических функций, и привязана для создания особым образом направленных перегрузок Картина еще осложнялась различным характером изменений при первом и втором испытаниях. Через 25 мин. после центрифугирования у двух собак происходило дальнейшее нарушение корковой деятельности по всем показателям: они не реагировали на сигналы, безусловные раздражители были вялыми, в конце опыта животные засыпали. У одной собаки во время подачи стереотипа раздражителей период возбуждения, судя по межсигнальным реакциям, сменялся глубоким разлитым торможением (сном). У трех животных на 25-й минуте после центрифугирования наблюдалось некоторое улучшение условнорефлекторной деятельности, скрытый период рефлексов уменьшался, собаки реагировали на большее, чем через 5 мин. после центрифугирования, число сигналов. Зато по другим показателям — реакциям на тормозные раздражители, наоборот, отмечалось ухудшение. После воздействия перегрузок нарушался характер самих двигательных реакций. Животные (особенно Малек и Пушинка) в ответ на условный раздражитель стали нажимать на педаль не 1–2 раза, как обычно, а несколько раз; причем нажимы быстро следовали один за другим. Движения отличались беспорядочностью, лапы двигались не вдоль педали, а поперек ее и выше, иногда по панели со световыми сигналами. Нередко подобные движения были неадекватными по своей силе (таких нарушений не наблюдалось только у Невы). Таблица 15. Число положительных условных реакций, дифференцировок и межсигнальных реакций у собак после воздействия перегрузок Изменения процессов, лежащих в основе реакций центральной нервной системы, очевидно, определялись характером сдвигов в области рефлекторной деятельности. Удлинение латентных периодов реакций и выпадение условных рефлексов в ответ на положительные раздражители свидетельствовали об ослаблении процесса возбуждения, а срывы дифференцировок и появление межсигнальных реакций — процесса торможения. О такой возможности в своем исследовании также говорит В.А. Кисляков (1956). У Невы при относительной сохранности возбудительного процесса нарушался процесс внутреннего торможения. У Малька наряду с повышенной возбужденностью отмечалась быстрая истощаемость нервных процессов. Возбуждение этой собаки носило разлитой характер и захватывало ряд разных анализаторов: животное реагировало не только на световые, по и на звуковые сигналы, реакции его были особенно хаотичными, сочетались со стремлением освободиться, издавать звуки и т. д. Реакции собак на отрицательные тормозные раздражители, появление межсигнальных реакций, хаотичное поведение — все свидетельствовало об ослаблении дифференцировочного, т. е. внутреннего торможения у многих животных. Однако эти же явления могли бы быть результатом развития в коре мозга пассивного разлитого торможения. Такое предположение подтверждалось вялостью некоторых собак. Значит, соотношение возбуждения и торможения изменялось в сторону преобладания торможения. Для всех животных без исключения было характерно раннее нарушение внутреннего процесса торможения (резкое сокращение правильных реакций на дифференцировочные раздражители, увеличение числа межсигнальных реакций и т. д.). И.П. Павлов считал внутреннее торможение наиболее хрупким и нежным видом тормозных реакций. Можно говорить об определяющей роли индивидуальных особенностей нервной системы собак. Нервная система Невы мало подвергалась влиянию перегрузок. У трех собак изменения по типу охранительного торможения наступали при первом обследовании, при втором у них отмечалась тенденция к восстановлению условнорефлекторных реакций. У Пушинки и Малька охранительное торможение развилось лишь к 25-й минуте после центрифугирования. Полной нормализации деятельности коры головного мозга в указанные сроки наблюдения ни у одного животного не наступило. Влияние на высшую нервную деятельность собак вибраций Вибрации даже в относительно небольших дозах, по мнению ряда исследователей, являлись сильным раздражителем для центральной нервной системы (Андреева-Галанина, 1956; Лукьянова, 1964; и др.). У собак действие местных вибраций с частотой 100 гц обусловливало повышение двигательных оборонительных рефлексов и растормаживание дифференцировки, а воздействия большей амплитуды — выпадение условных рефлексов. Кроме того, возникало снижение двигательных и секреторных оборонительных рефлексов (Михеева, 1955; Скачедуб, 1957). У крыс вибрация частотой 70 гц, амплитудой 0,4 м и длительностью 15 мин. снижала уровень условных рефлексов (Лившиц, Мейзеров, 1966). Делалась попытка установить механизмы, определяющие характер реакций центральной нервной системы в ответ на вибрационные воздействия. По мнению авторов ряда работ, вибрации вызывали значительную периферическую импульсацию от рецепторов кожи, мышц, суставов, вестибулярного аппарата. В низших отделах центральной нервной системы она обусловливала появление парабиотических фаз, а в коре — запредельного торможения. У животных со слабой нервной системой запредельное торможение было особенно ярко выраженным. Парабиотические явления в низших отделах центральной нервной системы в результате вибрационных воздействий были обнаружены Т.Н. Павловой (1958), М.Ф. Стома (1963, 1964), М.А. Кузнецовой, (1964, 1967). Л.Д. Лукьянова и Е.П. Казанская (1966) предполагали, что в основе изменений биоэлектрической активности головного мозга лежит возникновение генерализованного возбуждения, иррадиирующего в разные отделы головного мозга. Фазу генерального возбуждения, по мнению этих исследователей, сменяла фаза концентрации возбудительного процесса в зонах коры (в сенсомоторной, в зрительной). З.Н. Андреева (1964) развивала предположение, в соответствии с которым вибрационные воздействия вызывали внешнее торможение, а оно, в свою очередь, обусловливало появление растормаживания.  Рис. 28. Собака Нева, уложенная в специальный лоток, укреплена на вибростенде На артерию животного, выведенную в кожный лоскут, накладывается манжета для регистрации кровяного давления Рис. 28. Собака Нева, уложенная в специальный лоток, укреплена на вибростенде На артерию животного, выведенную в кожный лоскут, накладывается манжета для регистрации кровяного давления  Рис. 29. Собака Волхов во время вибровоздействия Рис. 29. Собака Волхов во время вибровоздействия В описанных ниже исследованиях, проведенных в плане отбора и подготовки животных, изучалось влияние вибрации на реакции центральной нервной системы собак. Характеристики вибраций соответствовали тем, которые предполагались при выведении космических кораблей на орбиту: 70 гц при амплитуде 0,4 мм, длительности 6 мин. (Георгиевский, Юганов, 1962). Использовался вибратор ВП-70. На рис. 28 и 29 показана подготовка и проведение этих экспериментов. Каждая собака подвергалась вибрированию от пяти до десяти раз. Опытам предшествовала тренировка собак, находящихся на станке вибростенда. Систему выработанных условных рефлексов проверяли непосредственно за 10–15 мин. до вибрирования, через 5–7 мин. после того, как выключался вибратор, и затем еще раз, спустя 20–25 мин. Таблица 16. Число условных реакций на положительные раздражители, дифференцировок и межсигнальных реакций у собак после воздействия вибраций Полученные данные (табл. 16) показали, что через 5 мин. после вибрирования у трех собак, относящихся к числу животных с сильной уравновешенной нервной системой, рефлекторная деятельность не изменялась или изменялась весьма незначительно: у двух из них наблюдалось два случая выпадения рефлексов и в одном случае растормаживалась дифференцировка, кроме того, отмечалось появление небольшого числа (четыре) межсигнальных реакций. На собаку Каму вибрация не влияла вообще. По прошествии 20 мин. реакции двух собак были изменены весьма незначительно: у них наблюдалось два случая выпадения рефлексов, растормаживалась одна дифференцировка, отмечалось появление небольшого числа (четыре) межсигнальных реакций. У остальных животных регистрировались более существенные сдвиги. Через 5 мин. две собаки не отреагировали на некоторые положительные сигналы, а через 25 мин. на 10 положительных раздражителей. Ухудшились тормозные функции, оказались сорваны некоторые дифференцировочные реакции, а еще через 20–25 мин. у одной собаки — все дифференцировки. Ослабление активного торможения через 7 и 20 мин. было видно также по большому количеству межсигнальных реакций. У одной собаки регистрировалось всего одно выпадение положительной реакции через 5 мин. и два через 20 мин., ухудшались реакции на дифференцировочные раздражители, появлялись межсигнальные реакции. У другого животного через 5–7 мин. после выключения вибратора нарушения были относительно незначительными, а через 20 мин. собака спала. Сравнивая реакции центральной нервной системы животных через 5 мин. после воздействия вибраций и через 20 мин., можно выделить собак, в условнорефлекторной системе которых за эти сроки сдвигов почти не произошло: это Кама и Нева. У одного животного к 20-й минуте по сравнению с 7-й минутой наблюдалось улучшение реакций центральной нервной системы. У двух на протяжении этого времени показатели высшей нервной деятельности значительно ухудшались: наблюдалась все прогрессирующая вялость. У одной собаки она сочеталась с периодами чрезмерно возбужденной активности. Мушка, несмотря на ухудшение многих реакций, не производила впечатления ни вялой, ни чрезмерно возбужденной. Известно, что некоторые исследователи (Минецкий, 1960; Стома, 1964) считали, что живые организмы не могут приспособиться к вибрации и давать менее выраженные реакции. Другие предполагали, что повторные воздействия снижали отрицательные сдвиги вибрирования, если вибрации были по своим параметрам не особенно значительными (Бутковская, 1951; Горбачевский, 1959). Эти данные заставляли предположить, что относительно хорошая переносимость вибраций тремя из шести собак могла объясняться их тренированностью. Для того, чтобы проверить такое предположение, были проведены опыты с собаками (Волна, Быстрый, Герта), ранее не подвергавшимися вибрации. Через 5–7 мин. после вибрирования у животных, как это видно по материалам табл. 16, были обнаружены значительные изменения условнорефлекторной деятельности: собаки часто производили хаотичные движения, появление которых не было связано с подачей сигналов, отсюда большое количество (24, 12 и 29) межсигнальных реакций. В то же время у них часто отсутствовали нажимы на педаль при подаче положительных сигналов. Некоторые дифференцировки были сорваны. Животные вели себя возбужденно: крутились в станке, издавали звуки, интенсивно ласкались к человеку и т. д. Через 25 мин. условнорефлекторная деятельность одной собаки несколько улучшилась, она стала вести себя значительно спокойнее: отреагировала на восемь сигналов, было сорвано три дифференцировки, на один дифференцировочный раздражитель она отреагировала правильно. Число межсигнальных реакций сократилось. У другой собаки через 20 мин. было отмечено три положительных реакции, все дифференцировки были сорваны, что, однако, при сильном сокращении реакций на сигнал не может свидетельствовать о внутреннем торможении. Число межсигнальных реакций равнялось четырем. В период обследования животное было малоподвижным и сразу после опыта заснуло. У собаки Быстрого через 20 мин. после центрифугирования наблюдалось почти полное восстановление условнорефлекторной деятельности. Данные этих опытов позволили предположить, что вибрация вызывала в коре головного мозга собак генерализованное возбуждение, что в литературе (Лукьянова, Казанская, 1966; Кузнецова, 1964) уже обсуждалось. В низших отделах центральной нервной системы импульсация с рецепторов кожи, мышц, суставов, вестибулярного аппарата обусловливала появление (Лившиц, Мейзеров, 1966) парабиотических фаз, а в коре — запредельного торможения. О запредельном торможении говорили данные по обследованию условнорефлекторной деятельности и других двух животных через 25 мин. после вибрирования. Следует отметить, что одна собака (Герта) при обследовании ее нервной системы была отнесена к числу животных со слабой нервной системой, а у таких животных запредельное торможение бывает особенно ярко выраженным. Влияние на реакции центральной нервной системы собак запусков в космос Изучение реакции центральной нервной системы в экстремальных условиях проводилось отечественной физиологией в работах школы И.П. Павлова. Внимание таким исследованиям уделяли и зарубежные исследователи (Hetherigton, 1959; и. др.). Однако их работы в плане влияния на рефлекторную деятельность обезьян космических полетов завершения не имели (Graybiel et al., 1960). В СССР изучалось влияние запусков животных (белых крыс) в космос на условнорефлекторную деятельность (Лукьянова, 1962). Исследования показали отсутствие существенных изменений высшей нервной деятельности. Собаки стоят на значительно более высокой ступени развития, чем грызуны, их мозг устроен более сложно, реакции существенно богаче, собаки способны воспринимать раздражения, на которые не реагируют крысы. Значит, правильно думать, что космические полеты могли иначе влиять на реакции центральной нервной системы и поведение собак. Далее описываются материалы, полученные при изучении реакций нервной системы животных после космических запусков. Исследовались четыре собаки. Пять животных по отношению к этим четырем собакам составляли контрольную группу: их возили на космодром и привозили обратно, а также подвергали всем другим процедурам. Проверка системы выработанных условных рефлексов проводилась за 1–4 дня до их отправки на космодром и в течение суток после приземления (они возвращались, поступали в виварий, их мыли, подвергали ветеринарному и медицинскому осмотру). Если у собак перед исследованием условнорефлекторной деятельности пищевое возбуждение было выражено недостаточно, их не кормили в течение 8 час, предшествующих эксперименту с подачей условных раздражителей. Трехкратная с интервалами в 10–20 мин. подача выработанного стереотипа раздражителей позволила высоко оценить условнорефлекторную деятельность собак, отобранных для запуска, а также контрольных животных. У всех собак не было отмечено даже минимальных нарушений условных реагирований (рис. 22 и 30). Изменения условнорефлекторной деятельности после запусков в космос показаны в табл. 17. Положительные реакции либо отсутствовали, либо имели место в относительно небольшом количестве случаев, которые к тому же преимущественно характеризовали рефлекторную деятельность Звездочки — одной из четырех собак, посланных в космос в кабине для человека. Отсутствие реакций на дифференцировочные раздражители было, очевидно, результатом не дифференцированного торможения, а могло объясняться отсутствием реакций вообще. Были обнаружены межсигнальные реакции, а также появление неполноценных (менее 15 г) нажимов, которые до полета не отмечались уже в течение многих месяцев. Все это говорило о существенном нарушении после космических полетов реакций центральной нервной системы этих собак. Контрольные животные, побывавшие на космодроме и доставленные в те же сроки в лабораторию, реагировали на 94,5% предъявляемых сигналов, в 90% случаев дифференцировки оказались ненарушенными. Величина латентного периода у них варьировала в пределах нормальной, межсигнальных реакций и неполноценных нажимов не было. Процесс изменения по мере подачи раздражителей стереотипа условнорефлекторной деятельности показан на рис. 31. Из рисунка видно, что у всех собак после полета во время подачи стереотипа раздражителей в одни периоды наблюдались активность, в другие — она почти полностью отсутствовала. Периоды активности были заполнены большим числом межсигнальных реакций, редкими реакциями на положительные раздражители, неоднократными срывами при дифференцировочных сигналах. Такая активность скорее была связана с возбужденностью собак, нежели с их деятельностью в ответ на стереотип раздражителей. Только в отношении Звездочки (см. рис. 31) можно было говорить о реакциях на предъявляемые воздействия.  Рис. 30. Условнорефлекторная деятельность собак до космических полетов А — Белка (12. V 1960 г.); Б — Жулька (13. XII 1966 г.); В — Звездочка (6. III 1961 г.). Обозначения те же, что на рис. 22, 23 Рис. 30. Условнорефлекторная деятельность собак до космических полетов А — Белка (12. V 1960 г.); Б — Жулька (13. XII 1966 г.); В — Звездочка (6. III 1961 г.). Обозначения те же, что на рис. 22, 23 У разных собак периоды активности имели неодинаковую степень выраженности и регистрировались на различных этапах подачи стереотипа условных раздражителей. У Белки и Стрелки активность имела интенсивную форму и протекала в две фазы. У Жульки наблюдался невыраженный цикл активности. Периоды активности длились от 1 до 3 мин., тогда как раздражители подавались в течение 7 мин., т. е. в основном собаки находились в пассивном состоянии. Таблица 17. Число положительных условных реакций, дифференцировок и межсигнальных реакций у собак через сутки после запуска в космос Таблица 17 (продолжение) Таблица 17 (продолжение)  Резкими были изменения в поведении собак. Стрелка в периоды активности тихонько скулила, в другие моменты лежала, часто отвернувшись от табло с раздражителями, не реагировала на щелчки аппарата, шум, производимый экспериментатором, и т. д. Белка в периоды неактивности стояла, смотрела на панель с сигнальными лампочками, реагировала на побочные раздражители, иногда начинала беспорядочно нажимать на крышки-педали, одно-двухразовые нажимы превращались в беспорядочные и торопливые движения, часто отмечались только взмахи лапой над педалями или нажимы с силой меньшей, чем 15 г.  Рис. 31. Условнорефлекторная деятельность собак после полета а — реакции на положительные сигналы; б — сила двигательной реакции собаки по планко-крышке менее 15 г; в — межсигнальные реакции; г, д — срыв дифференцировки А — Белка (12. VIII 1960 г.); В — Жулька (28. XII 1961 г.); В — Звездочка (27. III 1961 г.) Рис. 31. Условнорефлекторная деятельность собак после полета а — реакции на положительные сигналы; б — сила двигательной реакции собаки по планко-крышке менее 15 г; в — межсигнальные реакции; г, д — срыв дифференцировки А — Белка (12. VIII 1960 г.); В — Жулька (28. XII 1961 г.); В — Звездочка (27. III 1961 г.) Жулька вела себя крайне вяло, чаще всего лежала в расслабленной позе, а в конце эксперимента засыпала. Значительно была нарушена высшая нервная деятельность Звездочки; она реагировала только после 6-го раздражителя, латентный период был увеличен, дифференцировки оказались сорванными. Были зарегистрированы межсигнальные реакции, которых до этого у данного животного никогда не отмечалось. Собака вела себя значительно более вяло, чем обычно, нажимы на педали часто состояли из трех-четырех движений и иногда по силе были меньшими, чем в норме. На основании полученных данных можно сделать вывод о полном разрушении условнорефлекторной деятельности у трех собак и о значительном ее нарушении у четвертой. При этом можно предположить, что характерная для большинства животных смена периодов активности и пассивности свидетельствовала о цикличности процессов возбуждения и торможения в центральной нервной системе подопытных собак. Такое явление, возникающее под влиянием сильных раздражителей, отмечали и другие исследователи. Появление у Белки (см. рис. 31) большого числа межсигнальных реакций, увеличение движений и их хаотичность свидетельствовали о повышенной возбудимости корковых клеток и быстрой иррадиации в центральной нервной системе процесса возбуждения. Неактивное поведение (выпадение условнорефлекторных реакций и т. д.) говорило о торможении. Это позволяло предположить легкость возникновения возбуждения, так называемую его взрывчатость (И.П. Павлов), и быструю его истощаемость, о чем свидетельствовала резкая смена периодов хаотичной активности и полного отсутствия реагирований. Условнорефлекторная деятельность Стрелки обнаруживала взрывчатость возбуждения явно меньшую, чем у Белки, а истощаемость большую. Об этом говорило относительно небольшое время проявления возбуждения и длительные периоды отсутствия реакций. Процесс разрушения активного и появления запредельного торможения у этой собаки был более явным и глубоким: рефлексы в период неактивности не возникали, появление межсигнальных реакций и срывов дифференцировок сменялось чрезвычайной вялостью собаки. У Жульки истощенными оказались и процесс возбуждения, и процесс торможения. Процесс возбуждения выражался в появлении межсигнальных реакций, причем во втором цикле возбуждения их было только две, так что говорить о повышенной возбудимости этой собаки нет оснований. Ослабление процесса активного торможения подтверждалось хаотичностью реакций и теми же межсигнальными реакциями. Прогрессирующая вялость Жульки свидетельствовала о появлении запредельного торможения. У Звездочки изменения процесса возбуждения выражались в исчезновении прежней подвижности возбуждения: удлинялся латентный период, выпадали положительные реакции. Процесс активного торможения оказался нарушенным: животное неверно реагировало на большинство отрицательных раздражителей и у него, как и у других собак, наблюдались межсигнальные реакции. Таким образом, можно сделать вывод, что запуск в космос существенно повлиял на реакции центральной нервной системы подопытных собак. Нарушенными оказались процессы активного торможения. Выпадение условных рефлексов, увеличение латентного периода реакций (даже у Звездочки), вялость, сонливость говорят о развитии в нервной системе пассивного торможения типа запредельного. Выявленные изменения могли иметь характер функциональных сдвигов, возникавших в нервной системе как временное явление, а также быть выражением патологических особенностей, появившихся в результате космических полетов. В связи с этим целесообразно было исследовать процесс восстановления условнорефлекторной деятельности собак. Быстрое восстановление реакций животных на выработанный стереотип раздражений свидетельствовало бы о несущественности изменений реакций центральной нервной системы; медленное, но полное восстановление говорило бы о выраженности функциональных изменений. И, наконец, невозможность процесса восстановления условнорефлекторной деятельности позволила бы сделать вывод о патологических сдвигах. В связи с этим условнорефлекторная деятельность собак, побывавших в космосе, продолжала исследоваться в течение месяца после их приземления. Обследования проводились в 3, 7, 10, 15 и 20–25-е сутки 3 раза в день (с интервалами в 2–3 час). Данные, полученные по деятельности мозга Белки, Стрелки и Жульки в 3-й день после полета, не выявили никаких изменений по сравнению с материалами послеполетного обследования. Только у Звездочки наблюдалось некоторое улучшение выработанной системы. В связи с этим к большинству собак была применена система воздействий, направленная на восстановление разрушенного стереотипа условных рефлексов. Экспериментатор пробовал вызывать движения животных с помощью наталкивания, шлифовать их, применяя дифференцированную систему подкрепления, и т. д. (описано на с. 28 этой книги). Однако его попытки восстановить систему рефлексов к нужному эффекту не приводили. Условнорефлекторная деятельность Белки, Стрелки и Жульки на 7, 10, 15 и 20-й день после полета значительно не улучшилась. По-прежнему во время подачи стереотипа раздражителей в одни периоды наблюдалась активность, в другие она отсутствовала. Периоды активности были заполнены большим числом межсигнальных реакций, реакциями на некоторые положительные и дифференцировочные сигналы, временное соотношение периодов активности с периодами пассивности было примерно такое же, как сразу после полета. Все это заставляло сделать вывод об отсутствии улучшений в области реакций нервной системы, о взрывчатости и истощаемости возбуждения, о нарушениях активного торможения и о преобладании пассивного. У Звездочки, начиная с 8-го дня после полета (2. IV. 1961), показатели условнорефлекторной деятельности значительно улучшились, а в 10-й и 15-й день система выработанных рефлексов оказалась полностью восстановленной. Таким образом, отсутствие на протяжении полумесяца процесса восстановления условнорефлекторной деятельности Белки, Стрелки и Жульки позволяло сделать вывод о глубоких патологических сдвигах в реакциях центральной нервной системы этих собак под влиянием запуска в космос. Быстрое восстановление реакций Звездочки на стереотип раздражений свидетельствовало о менее существенных изменениях в области нервной системы этой собаки. Правомерно допустить, что причиной резких изменений реакций центральной нервной системы являлся комплекс секцусивных и симультанных раздражителей, связанных с полетом: разнообразные по силе, разнонаправленные ускорения, вибрации, шум, функциональные и даже физические нагрузки (сотрясение мозга при падении капсулы), мозговые гравитационно-вестибулярные и прочие влияния, невесомость. Они вносили существенный диссонанс в физиологические функции организма и прежде всего в реакции нервной системы, организующей первые формы адаптации к условиям среды. Обращает на себя внимание единая направленность изменений. Школой И.П. Павлова было доказано, что резкие и быстрые перестройки форм уравновешивания организма со средой вызывали так называемые сшибки — один из видов патологии нервных процессов. Очевидно, здесь имело место это явление. Известно, что характер расстройств нервной системы в значительной степени может зависеть от типологических особенностей собак. Однако, как об этом говорилось выше, например, Белка по целому ряду показателей была отнесена к сильному типу собак, тогда как у Звездочки за время определения особенностей ее рефлекторной деятельности фиксировались выпадения положительных условных рефлексов, срывы дифференцировок, появлялись межсигнальные реакции (см. рис. 23, 25, 26). Тем не менее система условных рефлексов Звездочки после полета восстановилась, а у Белки нет. Это заставляло думать об отсутствии в данном случае прямой связи между патологическим характером реакций собаки и типом ее высшей нервной деятельности. Можно предположить, что степень изменения состояния центральной нервной системы находилась в связи с устройством кабины космического корабля и другими условиями полета, например длительностью. Белка и Стрелка запускались в космос в кабинах для животных, являвшихся первыми образцами, и находились в полете трое суток. Запуск корабля с Жулькой был несколько позднее и также длился 3,5 суток. Звездочка летала меньше и в корабле типа «Восток», приспособленном для человека, т. е. в более комфортабельных условиях. Влияние запуска в космос на поведение и психическое состояние собак Поведение и психическое состояние собак может быть использовано как наглядный индикатор общего состояния живых организмов в необычных условиях и в том числе при полетах на космических кораблях, при освоении станций-лабораторий, при высадке на Луну и т. д. Попытки использования этих функций в качестве показателей степени влияния воздействий имели место при первых исследованиях жизнедеятельности собак в имитаторе кабины космического корабля (Газенко, Георгиевский, 1962), при полетах на кораблях-спутниках (Антипов и др., 1962; Журавлев, 1962). Говоря о поведенческих реакциях и о психике животных, надо отметить, с одной стороны, стойкую их сохранность (пищевого, оборонительного поведения, реагирования на человека), с другой — сильное изменение некоторых моментов поведения (выпадение сложных форм поведения, возникновение беспорядочных и лишних движений, а также малолабильных форм движений и позных состояний, появление скуления, стонов, озвученного дыхания). Характер этих изменений зависел от эмоционального состояния, сильного утомления, большого перенапряжения и определялся сдвигами в нервной деятельности и в психическом состоянии собак.  Рис. 32. Подготовка животных к полету Рис. 32. Подготовка животных к полету Вопросы поведения и психики животных экспериментально и теоретически разрабатывались еще на ранних стадиях развития биологической науки. Большим в этом отношении вкладом были разработанные в лабораториях И.П. Павлова объективные методические приемы: с их помощью определялось восприятие и другие психические функции животных (Орбели, 1961–1968; Зеленый, 1907; и др.). В последнее время были опубликованы экспериментально-теоретические работы по вопросам физиологии мозга, раскрывающие механизмы поведения и психические функции мозга. Несмотря на это, изучение поведения и психики животных продолжало оставаться нелегкой задачей. Это объяснялось сложностью самого объекта исследования, трудностями его расчленения, отсутствием четких объективных критериев поведенческих и психических реакций. Известную трудность представляли вопросы объективной регистрации поведения.  Рис. 33. В первые минуты после возвращения из космоса Рис. 33. В первые минуты после возвращения из космоса  Рис. 34. Собаки, побывавшие в космосе: Стрелка, Чернушка, Белка, Звездочка Рис. 34. Собаки, побывавшие в космосе: Стрелка, Чернушка, Белка, Звездочка В излагаемых ниже опытах была сделана попытка выработать у собак определенное поведение, охарактеризовать основные его моменты и по ним дать оценку психического состояния после орбитальных космических полетов. Эксперименты проводились на собаках Белке, Стрелке и Жульке. В качестве объекта изучения было выбрано поведение животных при перемещении — ходьбе, т. е. хорошо сложившаяся в филогенезе функция, вырабатываемая и автоматизирующаяся в онтогенезе. Она почти никогда не бывает полностью утраченной, но может существенно изменяться (темп движений, рисунок походки и т. д.). Перемещение изучалось с помощью снаряда: трехметровой деревянной доски шириною 13 см и двух тумб, в пазы которых вкладывались концы доски; бум располагался на разной высоте: 5, 45 и 125 см. В доску через каждые 25 см были вмонтированы катушки индукции. При перемещении собак, на одной из передних лап которых был привязан магнит, на ленте электрокардиографа возникали отметки. По этим отметкам определялся латентный период перемещений, скорость прохождения животным всего пути и его отрезков, задержки, участки бума, где собака соскакивала с доски, откуда она начала прыгать на тумбу, латентный период прыжка и т. д. Перемещение по доске вырабатывалось с помощью дрессировки по методу В.Л. Дурова (Дуров, 1924; Герд, 1957) в два этапа на маленькой высоте бума и на большой. В качестве раздражителей использовались наталкивающие натуральные воздействия, а затем искусственный сигнал. Подкрепляющими раздражителями были кусочки мяса или колбасы, подаваемые по дозированной системе (Герд, 1958). В результате собаки перемещались по буму в среднем через 5 сек. после подачи одного-двух искусственных сигналов. Животные шли по всей длине (300 см) доски (независимо от высоты). В 40–70 см от ее конца они прекращали движения и прыгали на вторую тумбу. Чтобы ответить на вопрос о том, как происходила ходьба на узкой доске, подошвы лап животных смазывались краской (Коган, 1969). Смещенной, суженной и расширенной считалась ходьба при условии отклонения животных от средней линии доски, а также при изменении ширины шага. В табл. 18 приведено время перемещения по буму разных собак (три последних занятия). Пульс, кровяное давление, дыхание и температура тела до запуска изменялись в пределах нормы. С помощью описанной методики делалась попытка путем исследования ряда поведенческих реакций оценить особенности психических функций, например восприятие и внимание, а также координацию движений. О функциях восприятия и внимания судили по скорости реагирования собак на искусственный сигнал, по скорости оценки ими расстояния для прыжка на тумбу, по количеству пробующих движений и т. д. Такие показатели, как потеря равновесия во время движения по доске, задержки, соскакивания с бума и, особенно в начале перемещения, неудавшиеся прыжки на тумбу, были показателями нарушений координации. Таблица 18 До запуска функции восприятия и внимания, а также координация движений собак, перемещающихся по доске, были нормальными, четко было сформировано сигнальное значение внешних раздражений. Собаки верно оценивали опорные возможности бума, расстояние для прыжка на вторую тумбу. Прыжки с тумбы на пол осуществлялись с правильных исходных позиций и предполагали мгновенную двигательную и сенсорную ориентировку. Во время проведения этой пробы животные выражали положительные эмоции: помахивали хвостом, осуществляли специфические движения с ориентацией на человека, спокойно вели себя во время перемещения, у них не было эмоциональной напряженности или вялости, при движении по доске отсутствовал страх. Реакции вегетативной системы были нормальными. Все это свидетельствовало о неудовлетворительном состоянии рецепторных функций и рабочих механизмов ходьбы и позволяло сделать вывод о хорошем состоянии отдельных поведенческих компонентов и психических реакций собак. После запуска поведенческие реакции собак при перемещении по доске на высоте 105 см, и это видно из табл. 19, значительно изменились. Животных было трудно заставить идти по доске. Экспериментатору приходилось много раз подавать сигналы, и собаки впервые вступили на доску только после четырех или пяти групп побуждающих попыток экспериментатора. Еще больше усилий требовалось для того, чтобы заставить их идти по доске. Сделав один-два шага, они стремились повернуть назад, а четыре-пять — спрыгнуть с доски. Все это говорило о том, что животные заранее оценивали эти действия как неблагоприятные, и такая оценка, судя по начальным шагам и прыжкам (часто прыжкам-падениям), соответствовала действительной оценке возможностей собак. Перемещения до конца доски впервые были получены у Белки после 11 групп повторных попыток, у Жульки — после 25 и у Звездочки — после 13. Поведенческие реакции этих животных были различными. Белка перемещалась по доске быстро, без задержек и пробующих движений. Значительные отклонения лап от средней линии перемещения, их постановка на самый край и скольжение, другие виды потери равновесия преодолевались животным в ходе движения, зачастую путем увеличения скорости и увеличения двигательных ошибок. Создавалось впечатление, что если бы не такая скорость, животное не могло бы перемещаться по доске. Таблица 19. Характеристика перемещения собак по доске до и после запуска в космос Жульку, наоборот, характеризовало медленное движение, большое количество задержек, пробующих движений. В связи с этим количество потерь равновесия у этой собаки было меньше, чем у Белки. Передвижение по доске и прыжки с нее часто совершались без необходимой предварительной ориентировки и с неправильных позиций, собаки долго примеривались, а касаясь пола, плохо пружинили тело. Все это говорило об ухудшении координации движений. Об ухудшении восприятия и понижении внимания у Белки говорили двигательные ошибки, связанные с неверной оценкой ширины доски, расстояний и т. д., часто они кончались потерями равновесия, а иногда и падениями. Те же явления имели место и у двух других собак. Интересным феноменом было появление движений, которые не наблюдались до запуска, а после запуска у Жульки, Стрелки и Звездочки получили широкое распространение. Условно такие действия были названы пробными. Собаки, находясь на доске, делали попытку поставить на очередную позицию переднюю лапу, но, еще не начав перемещать на нее центр тяжести, переставляли эту лапу в исходное положение, а затем в другое место. Иногда пробные движения имели еще менее завершенную форму: животные проводили лапой или дотрагивались ею до поверхности доски и тут же убирали ее снова. В ряде случаев они ограничивались подобным движением, не касаясь доски. Все это свидетельствовало о том, что после запуска собаки не могли в оценке опорных поверхностей ограничиваться показаниями зрительного анализатора и в наиболее ответственные моменты привлекали также и другие виды ориентировки. Эмоциональное состояние собак во время этих проб после космических полетов было неудовлетворительным. Отмечалась одеревенелость мускулатуры, в некоторых случаях дрожь, что говорило о страхе. Помахивания хвостом не регистрировались. Были случаи непроизвольных мочеиспусканий и дефекации. После перемещения по буму животные часто не реагировали на многие раздражители. Иногда дышали открытой пастью, часто и поверхностно, иногда ложились и почти не двигались. Все это производило впечатление сильной утомленности, совершенно не регистрируемой до запуска в космос. Для оценки психического состояния собак также были использованы эмоциональные пробы. Полученные при этом данные, сведенные в табл. 20, говорят о значительном неблагоприятном изменении вегетативных реакций. Пробы, очевидно, вызывали в психике собак существенные сдвиги, не типичные для их нормального состояния. Движения были скованы, мускулатура напряжена, отмечалась дрожь тела, скуление, непроизвольное выделение мочи, вялость, ненормальная расслабленность, неподвижность или патологическая торопливость. Таблица 20. Частота пульса и дыхания животных, время удерживания груза при эмоциональных пробах на 2–3-й день после запуска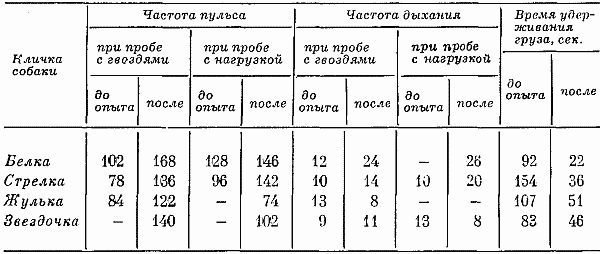 Судя по сокращению времени удерживания груза и по ярко выраженным отрицательным поведенческим реакциям при нагрузках (скуление, попытки лечь на гвозди, прислониться к стенке и т. д.), собаки после запуска в космос стали быстро утомляться. Таким образом, полученные факты говорят о значительном ухудшении ряда поведенческих реакций собак, связанных с их восприятием, эмоциональным состоянием, координацией движений. Значит, запуск в космос влиял на поведение и психическую деятельность собак. Сопоставление этих данных с данными, приведенными в предыдущем разделе, позволяет говорить о соответствии материалов, характеризующих поведение животных, материалам, определяющим другие изменения. Поведение собак отражало характер тех сдвигов, которые были зарегистрированы при анализе реакций центральной нервной системы. Возникновение движений ходьбы и быстрое их прекращение, замедленность и вялость реакций собак, по-видимому, объяснялись ухудшением у них процессов возбуждения, их взрывчатостью и быстрой истощаемостью, а также нарушением процессов активного торможения и, наоборот, развитием пассивного. Об ухудшении активных видов торможения говорит неудовлетворительная ориентировка животных на доске и неточность их движений, о развитии пассивных движений — быстрая утомляемость и сонливость. Интересно, что в приведенных нами фактах отчетливо прослеживались и более детальные характеристики поведения, определяемого сдвигами в центральной нервной системе. Так, например, у Белки была обнаружена повышенная возбудимость, большая, чем у Жульки и Стрелки, и соответственно этому поведение этой собаки характеризовалось более быстрым появлением неточных реакций, скорыми и торопливыми движениями. Наоборот, факты, подтверждающие связь характера изменения поведения с типом высшей нервной деятельности, в наших опытах не были обнаружены. Белка, да и Жулька, во время опытов, направленных на выявление особенностей их высшей нервной деятельности, были отнесены к числу собак с сильной нервной системой, и тем не менее после запуска их поведение изменялось более значительно, чем, например, у Звездочки, у которой ни процесс возбуждения, ни процесс торможения во время предварительных обследований не обладали такой силой, как у Белки и Стрелки. Особо следует остановиться на резком ухудшении эмоционального состояния собак после запуска. Выше было показано, что угнетенность психики у некоторых собак возникает после их длительного пребывания в камере. Здесь тот же эффект в его яркой форме был достигнут после запуска в космос. Поскольку мы наблюдали его в сочетании со сдвигами в центральной нервной системе, а эти сдвиги, как доказано в предыдущем разделе, были характерны для животных, использованных в полетах, то ухудшение психического состояния следовало признать результатом значительных сдвигов в высшей нервной деятельности собак. Очевидно, при резких неблагоприятных изменениях в области возбудительных и тормозных процессов, описанные изменения психических процессов обусловливают чрезмерно осторожное и неактивное поведение животных и тем самым позволяют избегать сильных раздражителей. В этом заключается, по-видимому, один из механизмов адаптации собак к состоянию их центральной нервной системы. |
|
||